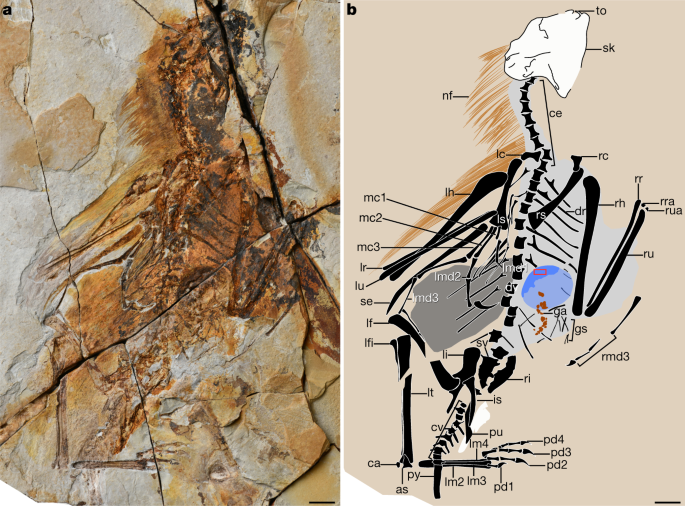
Summary
Powered flight superior independently in vertebrates in the pterosaurs, birds and bats, each of which has a numerous configuration of the bony ingredients and epidermal constructions that originate the wings1,2. Whereas the early fossil records of pterosaurs and bats are sparse, mounting proof (primarily from China) of feathered non-avian dinosaurs and stemward avians that earn primarily from the Center–Upper Jurassic and Decrease Cretaceous courses has enabled the sluggish piecing collectively of the origins of avian flight3,4. These fossils portray that, terminate to the foundation of flight, dinosaurs closely associated to birds were experimenting with a vary of cruise constructions3,5. Practical one of essentially the most surprising of these is that of the scansoriopterygid (Theropoda, Maniraptora) Yi qi, which has membranous wings—a flight apparatus that changed into beforehand unknown amongst theropods however that’s normal by both the pterosaur and bat lineages6. This observation changed into no longer universally current7. Here we describe a newly identified scansoriopterygid—which we name Ambopteryx longibrachium, gen. et sp. nov.—from the Upper Jurassic duration. This specimen presents strengthen for the long-established existence of membranous wings and the styliform ingredient in the Scansoriopterygidae, as successfully as proof for the diet of this enigmatic theropod clade. Our analyses portray that marked adjustments in cruise structure superior terminate to the crash up between the Scansoriopterygidae and the avian lineage, because the 2 clades travelled alongside very numerous paths to turning into volant. The membranous wings supported by elongate forelimbs which may perchance perchance well be most long-established in scansoriopterygids almost definitely record a quick-lived experimentation with volant behaviour, and feathered wings were in a roundabout method favoured throughout the later evolution of Paraves.
Access alternate ideasAccess alternate ideas
Subscribe to Journal
Win beefy journal receive admission to for 1 year
227,39 €
supreme 4,46 € per self-discipline
All costs encompass VAT for Germany.
Rent or Grab article
Win time miniature or beefy article receive admission to on ReadCube.
from$8.99
All costs are NET costs.
Data availability
All data—alongside with the measurements, provide data for morphometric prognosis and phylogenetic data matrix—that strengthen the findings of this research are included as Supplementary Data. The specimen (IVPP V24192) described on this understand is archived and on hand on search data from from the IVPP. A Life science Identifier for the newly described species has been registered at ZooBank (http://zoobank.org/): urn:lsid:zoobank.org:act:0A2DE2F0-CE78-4149-B0BD-A0DE91FC1328. Any numerous associated data are on hand from the corresponding creator upon realistic search data from.
Further data
Writer’s articulate: Springer Nature stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.
Gatesy, S. M. & Middleton, K. M. in Fins Into Limbs: Evolution, Model, and Transformation (ed. Hall, B. K.) 269–283 (Univ. Chicago Press, Chicago, 2007).
-
- 2.
Norberg, U. Vertebrate Flight: Mechanics, Physiology, Morphology, Ecology and Evolution (Springer, Berlin, 1990).
-
- 3.
Xu, X. et al. An integrative technique to conception chook origins. science 346, 1253293 (2014).
- 4.
Gatesy, S. M. & Middleton, K. M. Bipedalism, flight, and the evolution of theropod locomotor vary. J. Vertebr. Paleontol. 17, 308–329 (1997).
- 5.
Chatterjee, S. & Templin, R. J. Biplane cruise planform and flight performance of the feathered dinosaur Microraptor gui. Proc. Natl Acad. Sci. USA 104, 1576–1580 (2007).
- 6.
Xu, X. et al. A extraordinary Jurassic maniraptoran theropod with preserved proof of membranous wings. Nature 521, 70–73 (2015).
- 7.
Padian, K. Dinosaur up in the air. Nature 521, 40–41 (2015).
- 8.
Liu, Y., Liu, Y., Ji, S. A. & Yang, Z. U–Pb zircon age for the Daohugou Biota at Ningcheng of Interior Mongolia and comments on associated elements. Chin. Sci. Bull. 51, 2634–2644 (2006).
- 9.
Huang, D. Yanliao biota and Yanshan circulation (in Chinese language). Acta Palaeontologica Sin. 54, 501–546 (2015).
- 10.
Campione, N. E., Evans, D. C., Brown, C. M. & Carrano, M. T. Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions. Methods Ecol. Evol. 5, 913–923 (2014).
- 11.
Other folks, W. S., Currie, P. J. & Norell, M. A. Oviraptorosaur tail forms and positive aspects. Acta Palaeontol. Pol. 59, 553–567 (2013).
- 12.
O’Connor, J. K. & Sullivan, C. Reinterpretation of the Early Cretaceous maniraptoran (Dinosauria: Theropoda) Zhongornis haoae as a scansoriopterygid-esteem non-avian, and morphological resemblances between scansoriopterygids and basal oviraptorosaurs. Vertebr. Palasiat. 52, 3–30 (2014).
- 13.
Gatesy, S. M. & Thomason, J. in Functional Morphology in Vertebrate Paleontology (ed. Thomason, J. J.) 219–234 (Cambridge Univ. Press, Cambridge, 1995).
-
- 14.
Zhang, F., Zhou, Z., Xu, X., Wang, X. & Sullivan, C. A extraordinary Jurassic maniraptoran from China with elongate ribbon-esteem feathers. Nature 455, 1105–1108 (2008).
- 15.
Zhang, F., Zhou, Z., Xu, X. & Wang, X. A juvenile coelurosaurian theropod from China signifies arboreal habits. Naturwissenschaften 89, 394–398 (2002).
- 16.
Turner, A. H., Makovicky, P. J. & Norell, M. A. A review of dromaeosaurid systematics and paravian phylogeny. Bull. Am. Mus. Nat. Hist. 371, 1–206 (2012).
- 17.
Xu, X. et al. A brand unique feathered maniraptoran dinosaur fossil that fills a morphological gap in avian foundation. Chin. Sci. Bull. 54, 430–435 (2009).
- 18.
Balanoff, A. M. & Norell, M. A. Osteology of Khaan mckennai (Oviraptorosauria: Theropoda). Bull. Am. Mus. Nat. Hist. 372, 1–77 (2012).
- 19.
Hutchinson, J. R. The evolution of pelvic osteology and soft tissues on the dual carriageway to extant birds (Neornithes). Zool. J. Linn. Soc. 131, 123–168 (2001).
- 20.
Zhang, F. et al. Fossilized melanosomes and the shade of Cretaceous dinosaurs and birds. Nature 463, 1075–1078 (2010).
- 21.
Thorington, J. R. W., Darrow, K. & Anderson, C. G. Hover tip anatomy and aerodynamics in flying squirrels. J. Mamm. 79, 245–250 (1998).
- 22.
Oshida, T., Hiraga, H., Nojima, T. & Yoshida, M. C. Anatomical and histological notes on the foundation of the long accessory styliform cartilage of the Russian flying squirrel, Pteromys volans orii. Mammal Sight 25, 41–48 (2000).
- 23.
Dial, K. P. Hover-assisted incline working and the evolution of flight. science 299, 402–404 (2003).
- 24.
Lovette, I. J. & Fitzpatrick, J. W. Manual of Bird Biology third edn (John Wiley & Sons, Hoboken, 2016).
-
- 25.
Sullivan, C. et al. The vertebrates of the Jurassic Daohugou biota of northeastern China. J. Vertebr. Paleontol. 34, 243–280 (2014).
- 26.
Liu, Y., Liu, Y. & Zhang, H. LA-ICPMS zircon U-Pb relationship in the Jurassic Daohugou beds and correlative strata in Ningcheng of Interior Mongolia. Acta Geol. Sin. 80, 733–742 (2006).
- 27.
Liu, Y.-Q. et al. Timing of the earliest identified feathered dinosaurs and transitional pterosaurs older than the Jehol biota. Palaeogeogr. Palaeoclimatol. Palaeoecol. 323–325, 1–12 (2012).
- 28.
Chu, Z. et al. High-precision U-Pb geochronology of the Jurassic Yanliao biota from Jianchang (western Liaoning Province, China): age constraints on the upward thrust of feathered dinosaurs and eutherian mammals. Geochem. Geophys. Geosyst. 17, 3983–3992 (2016).
- 29.
Xu, X., Zhou, Z., Sullivan, C., Wang, Y. & Ren, D. An up to this level review of the Center–Late Jurassic Yanliao Biota: chronology, taphonomy, paleontology and paleoecology. Acta Geol. Sin. 90, 2229–2243 (2016).
- 30.
Benson, R. B. J. et al. Charges of dinosaur body mass evolution articulate 170 million years of sustained ecological innovation on the avian stem lineage. PLoS Biol. 12, e1001853 (2014).
- 31.
Christiansen, P. & Fariña, R. A. Mass prediction in theropod dinosaurs. Hist. Biol. 16, 85–92 (2004).
- 32.
Serrano, F. J., Palmqvist, P. & Sanz, J. L. Multivariate prognosis of neognath skeletal measurements: implications for body mass estimation in Mesozoic birds. Zool. J. Linn. Soc. 173, 929–955 (2015).
- 33.
Campione, N. E. & Evans, D. C. A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods. BMC Biol. 10, 60 (2012).
- 34.
Goloboff, P. A. & Catalano, S. A. TNT version 1.5, alongside with a beefy implementation of phylogenetic morphometrics. Cladistics 32, 221–238 (2016).
- 35.
Brusatte, S. L. in Computational Paleontology (ed. Elewa, A. M. T.) 53–74 (Springer, Heidelberg, 2011).
-
- 36.
Laurin, M. The evolution of body dimension, Cope’s rule and the foundation of amniotes. Syst. Biol. 53, 594–622 (2004).
- 37.
Wang, M. & Lloyd, G. T. Charges of morphological evolution are heterogeneous in Early Cretaceous birds. Proc. R. Soc. Lond. B 283, 20160214 (2016).
- 38.
Bapst, D. W. paleotree: an R equipment for paleontological and phylogenetic analyses of evolution. Methods Ecol. Evol. 3, 803–807 (2012).
- 39.
Revell, L. J. Size-correction and necessary ingredients for interspecific comparative reviews. Evolution 63, 3258–3268 (2009).
- 40.
Benson, R. B. J. & Choiniere, J. N. Charges of dinosaur limb evolution present proof for distinctive radiation in Mesozoic birds. Proc. R. Soc. Lond. B 280, 20131780 (2013).
- 41.
Hu, D. et al. A bony-crested Jurassic dinosaur with proof of iridescent plumage highlights complexity in early paravian evolution. Nat. Commun. 9, 217 (2018).
- 42.
Paradis, E., Claude, J. & Strimmer, K. APE: analyses of phylogenetics and evolution in R language. Bioinformatics. 20, 289–290 (2004).
- 43.
Felsenstein, J. Phylogenies and the comparative contrivance. Am. Nat. 125, 1–15 (1985).
- 44.
Revell, L. J. phytools: an R equipment for phylogenetic comparative biology (and numerous issues). Methods Ecol. Evol. 3, 217–223 (2012).
Acknowledgements
We thank S.-X. Jiang, D.-Y. Huang, Y.-H. Pan and Z.-Q. Yu for dialogue, Q.-R. Meng for serve in the self-discipline, T. Zhao for taking scanning electron microscopy photos, D.-H. Li for specimen preparation and W. Gao for photographing. This research changed into supported by the National Pure science Foundation of China (41688103; 41722202), Formative years Innovation Promotion Affiliation CAS (2016073) and the Recount Key Laboratory of Lithospheric Evolution (Z201604).
Reviewer data
Nature thanks Thomas Richard Holtz, Peter Makovicky and Kevin Padian for his or her contribution to the peer review of this work.
Prolonged data figures and tables
-
Prolonged Data Fig. 1 Further photos of Ambopteryx, IVPP V24192.
a, Counter slab. b, Skeletal reconstruction in step with preserved bones. c, Skull. d, Gastroliths and the unidentified bony abdomen snort. Abbreviations as in Fig. 1, with the exception of for: fe, feather associated to the neck; lil, left ilium;; lti, left tibia; pd, pedal digits; and ub, unidentified bony ingredient. The white field signifies the status from which the pattern changed into taken for histological prognosis. Scale bars, 10 mm (a, c, d), 20 mm (b).
-
Prolonged Data Fig. 2 Bone histology of Ambopteryx.
a–d, Thin substandard-part of the left humerus (a, b) and the unidentified bony abdomen snort (c, d). The arrowheads articulate the osteocyte lacunae. Scale bars, 100 μm (a–c), 200 μm (d).
-
Prolonged Data Fig. 3 Anatomy of Ambopteryx, IVPP V24192.
a–d, Interpretative drawings of the neck and pectoral girdle (a), caudal vertebrae and pygostyle (b), left forelimb (c) and pelvis and left hindlimb (d). Abbreviations as in Figs. 1, 2, with the exception of for: ca, caudal vertebrae; cm, calcaneum; de, deltopectoral crest of humerus; invent, dorsal vertebrae; I–III, metacarpals I–III; ip, iliac peduncle of ilium; mtII–IV, metatarsals II–IV; p1–4, pedal digits I to IV; and st, styliform ingredient. Scale bars, 10 mm (a–d).
-
Prolonged Data Fig. 4 Forelimb comparisons between Ambopteryx and Yi.
a, Left forelimb of Ambopteryx (IVPP V24192). b–d, Left forelimb (b), honest humerus (c) and the honest styliform ingredient (d) of Yi (STM 31-2). The proximal margins of the humeri are marked in white dashed lines to portray the diversifications between these two taxa. Abbreviations as in Figs. 1, 2, with the exception of for: st, styliform ingredient. Scale bars, 10 mm (a), 20 mm (b–d).
-
Prolonged Data Fig. 5 Comparisons of hand morphology amongst Scansoriopterygidae.
a–c, Line drawings of the fingers of Ambopteryx (a), Epidendrosaurus (b) and Yi (c). Scale bars, 10 mm (a, b), 20 mm (c).
-
Prolonged Data Fig. 6 Scanning electron microscopy photos of the soft tissues which may perchance perchance well be preserved in Ambopteryx.
a–d, Feather samples associated to the neck. e–f, Samples of membranous tissues taken from the self-discipline between the left femur and left handbook digits. Arrows denote the positions of the samples. Scale bars, 2 μm.
-
Prolonged Data Fig. 7 Time-scaled recovered strict-consensus tree of Mesozoic coelurosaurians.
Bremer and bootstrap values are labelled terminate to the corresponding node in heroic italic and appropriate non-heroic font, respectively.
-
Prolonged Data Fig. 8 Compiled huge-tree of the sampled Mesozoic coelurosaurians normal in morphometric analyses.
Total tree for the coelurosaurians normal in generating the PPCA morphospaces confirmed in Fig. 3b, c.
Supplementary data
-
Supplementary Data
This file accommodates Supplementary Text Sections 1–5; which encompass extra anatomical description, abdomen contents and diet of Ambopteryx longibrachium, Supplementary Tables 1, 2, 4 and 5, and the information normal in the phylogenetic prognosis.
-
Reporting Summary
-
Supplementary Desk 3
This file accommodates appendicular limb bone measurements of Mesozoic coelurosaurians normal in the phylogenetic necessary ingredients prognosis.
Rights and permissions
To accumulate permission to re-use snort from this text talk to RightsLink.
Comments
By submitting a observation you compromise to abide by our Terms and Neighborhood Pointers. Even as you receive something abusive or that would now not conform to our phrases or guidelines please flag it as flawed.



Leave a comment
Sign in to post your comment or sign-up if you don't have any account.